Jedná se o schopnost pufrovacího systému působit proti změnám pH prostředí.
Nazývá se rozsah hodnot pH, nad a pod kterým přestává pufrační účinek nárazníková zóna.
Je rovnocenná pH = pK ± 1
Pufrovací kapacita (B) je vyjádřena jako počet molárních ekvivalentů silné kyseliny nebo zásady, které je nutné přidat do jednoho litru pufru, aby se pH posunulo o jednu.
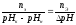
B – vyrovnávací kapacita,
nЭ – množství molárního ekvivalentu silné kyseliny nebo zásady,
pHН – počáteční hodnota pH (před přidáním kyseliny nebo zásady)
pHК – konečná hodnota pH (po přidání kyseliny nebo zásady)
V praxi se vyrovnávací kapacita vypočítá pomocí vzorce:
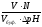
V – objem kyseliny nebo zásady,
N – ekvivalentní koncentrace kyseliny nebo zásady,
Vfanoušek.– objem tlumivého roztoku,
Následující údaje naznačují, že působí proti změnám pH krve. Chcete-li posunout pH krve o jednu jednotku do alkalické oblasti, musíte do krve přidat 70krát více NaOH než do stejného objemu čisté vody. Pro změnu pH o jednu na kyselou oblast by mělo být do krve přidáno 320krát více kyseliny chlorovodíkové než do stejného objemu čisté vody.
Kapacita pufru závisí na koncentraci elektrolytů a poměru složek pufru. Roztoky s vyšší koncentrací složek a poměrem složek rovným jedné mají největší pufrační kapacitu.
Pufrovací kapacita arteriální krve je 25,3 mmol/l, venózní krve 24,3 mmol/l, sliny mají pufrační kapacitu a jsou určeny bikarbonátovým, fosfátovým a proteinovým systémem. Tlumicí kapacita slin se mění pod vlivem řady faktorů: sacharidová dieta snižuje tlumivou kapacitu slin, vysokoproteinová dieta ji zvyšuje. Výskyt zubního kazu je menší u jedinců s vysokou pufrovací kapacitou.
V lidském těle působí proteinové, hemoglobinové, fosfátové a bikarbonátové pufry.
Tlumivé systémy těla.
Tvoří 53 % kapacity vyrovnávací paměti a je reprezentován:
Bikarbonátový pufr je hlavním pufrovacím systémem krevní plazmy; je to systém rychlé reakce, protože je produktem jeho interakce s kyselinami CO2 – rychle se vylučuje plícemi. Kromě plazmy se tento pufrový systém nachází v červených krvinkách, intersticiální tekutině a ledvinové tkáni.
Mechanismus působení.
Pokud se kyseliny hromadí v krvi, množství HCO klesá3 – a dojde k reakci: NSO3 – + N + ↔ N2S3 ↔ N2O + CO2↑. Přebytek je odstraněn plícemi. Hodnota pH krve však zůstává konstantní, protože objem plicní ventilace se zvyšuje, což vede ke snížení objemu CO2
Se zvýšením alkality krve, koncentrace HCO3 – zvyšuje: N2S3 + OH – ↔ NSO3 – + N2O.
To vede k pomalejší ventilaci plic, takže CO2 se hromadí v těle a poměr pufrů zůstává nezměněn.
Představuje 35 % kapacity vyrovnávací paměti.
Hlavní pufrovací systém červených krvinek, který tvoří asi 75 % celkové pufrační kapacity krve. Účast hemoglobinu na regulaci pH krve je spojena s jeho úlohou v transportu kyslíku a CO2. Systém hemoglobinového pufru v krvi hraje významnou roli v několika fyziologických procesech najednou: dýchání, transport kyslíku v tkáních a udržování konstantního pH uvnitř červených krvinek a nakonec v krvi.
Je reprezentován dvěma slabými kyselinami – hemoglobinem a oxyhemoglobinem a jejich konjugovanými bázemi – hemoglobinátovými a oxyhemoglobinátovými ionty:
Oxyhemoglobin je silnější kyselina (pKа = 6,95) než hemoglobin (pKа = 8,2). Při pH = 7,25 (uvnitř červených krvinek) je oxyhemoglobin ionizován o 65 % a hemoglobin o 10 %, takže přidání kyslíku k hemoglobinu snižuje hodnotu pH krve, protože se tvoří silnější kyselina. Na druhou stranu, jak se z oxyhemoglobinu ve tkáních uvolňuje kyslík, hodnota pH krve se opět zvyšuje.
Tlumivé vlastnosti HHb jsou primárně dány možností interakce kyselino-reaktivních sloučenin s draselnou solí hemoglobinu za vzniku ekvivalentního množství odpovídající draselné soli kyseliny a volného hemoglobinu:
Výsledný hydrogenuhličitan (HCO3) vyrovnává množství příchozích H2S3pH se udržuje, když se potenciální molekuly H disociují2S3 a výsledné hemoglobinové kyseliny.
Takto se pH krve udržuje v normálních mezích i přes vstup obrovského množství CO do žilní krve2 a další kysele reaktivní metabolické produkty.
V kapilárách plic hemoglobin (HHb) absorbuje kyslík a mění se na HHbO2, což vede k určitému okyselení krve, vytěsnění určitého množství H2S3 z bikarbonátů a snížení alkalické rezervy krve a v tkáních ji odevzdává a absorbuje CO2.
Kromě toho je hemoglobinový pufr komplexní protein a působí jako proteinový pufr.
Tvoří 5 % kapacity vyrovnávací paměti. Obsaženo jak v krvi, tak v buněčné tekutině jiných tkání, zejména ledvin. V buňkách je reprezentován K solemi2NRA4 a KN2RO4a v krevní plazmě a mezibuněčné tekutině Na2HPO4 a NaH2PO4. Funguje hlavně v plazmě a zahrnuje: dihydrogenfosfátový iont H2RO4 – a hydrogenfosforečnanový ion HPO4 2-.
Poměr [HPO4 2-]/[H2PO4 – ] v krevní plazmě (při pH = 7,4) se rovná 4:1. V důsledku toho má tento systém pufrační kapacitu pro kyselinu větší než pro zásadu.
Například při zvýšení koncentrace kationtů H + v intracelulární tekutině, například v důsledku zpracování masité potravy, jsou neutralizovány ionty HPO4 2-:
Vzniklý nadbytek dihydrogenfosforečnanu je vylučován ledvinami, což vede ke snížení pH moči.
Když se koncentrace zásad v těle zvýší, například při konzumaci rostlinné stravy, jsou neutralizovány H ionty2RO4 1-:
Vzniklý nadbytek hydrogenfosforečnanu je vylučován ledvinami a zvyšuje se pH moči.
Vylučování určitých složek fosfátového pufrového systému močí v závislosti na zpracovávané potravině vysvětluje široký rozsah hodnot pH moči – od 4,8 do 7,5. Fosfátový pufrovací systém krve se vyznačuje nižší pufrovací kapacitou než bikarbonátový systém v důsledku nízké koncentrace krevních složek. Tento systém však hraje rozhodující roli nejen v moči, ale i v dalších biologických prostředích – v buňce, ve šťávách trávicích žláz, v moči.
Tvoří 5 % kapacity vyrovnávací paměti. Skládá se z bílkovinné kyseliny a její soli tvořené silnou zásadou.
Pt – COOH – protein-kyselina
Pt – COONa – protein-sůl
Když se v těle tvoří silné kyseliny, interagují s proteinovou solí. Tím vzniká ekvivalentní množství proteinové kyseliny: HC1 + Pt-COONa ↔ Pt-COOH + NaCl. Podle zřeďovacího zákona W. Ostwalda zvýšení koncentrace slabého elektrolytu snižuje jeho disociaci, pH zůstává prakticky nezměněno.
Jak alkalické potraviny přibývají, interagují s
Pt-COOH: NaOH + Pt-COOH ↔ Pt-COONa + H2O
Množství kyseliny klesá. Koncentrace H+ iontů se však zvyšuje v důsledku potenciální kyselosti proteinové kyseliny. pH se tedy prakticky nemění.
Protein je amfoterní elektrolyt, a proto vykazuje svůj vlastní pufrovací účinek.
Podívejme se na interakci nárazníkových systémů v těle ve fázích:
Během výměny plynů v plicích vstupuje kyslík do červených krvinek, kde probíhá reakce:
Při pohybu krve do periferních částí oběhového systému se z ionizované formy HbO uvolňuje kyslík2 –
V tomto případě se krev mění z arteriální na venózní. Kyslík uvolněný ve tkáních se spotřebuje na oxidaci různých substrátů, což má za následek tvorbu CO2, z nichž většina vstupuje do erytrocytů.
V erytrocytech v přítomnosti karboanhydrázy dochází významnou rychlostí k následující reakci:
Výsledné přebytečné protony se vážou na hemoglobinátové ionty:
Vazbou protonů se posouvá rovnováha reakce stupně (3) doprava, v důsledku toho se zvyšuje koncentrace hydrogenuhličitanových iontů a ty difundují membránou do plazmatu. V důsledku protidifúze iontů lišících se acidobazickými vlastnostmi (chloridový iont je protolyticky neaktivní, hydrogenuhličitanový iont je v tělesných podmínkách zásadou) hydrokarbonát-chloridový posun. To vysvětluje kyselejší reakci prostředí v erytrocytech (pH = 7,25) ve srovnání s plazmou (pH = 7,4).
Bikarbonátové ionty vstupující do plazmy neutralizují přebytečné protony, které se tam hromadí v důsledku metabolických procesů:
Výsledný CO2 interaguje se složkami proteinového pufrovacího systému:
S2 + Рt-NH2 ↔ Pt-NHCOOH ↔ H + + Pt-NHCOO –
Přebytečné protony jsou neutralizovány fosfátovým pufrem:
Po návratu krve do plic se v ní zvyšuje koncentrace oxyhemoglobinu (1. stupeň), který reaguje s hydrogenuhličitanovými ionty, které nedifundovaly do plazmy:
Výsledný CO2 se vylučuje plícemi. V důsledku poklesu koncentrace HCO3 – jsou pozorovány ionty v této části krevního řečiště, jejich difúze do erytrocytů a difúze chloridových iontů v opačném směru.
Ledviny také hromadí přebytečné protony v důsledku reakce:
který je neutralizován hydrofosfátovými ionty a amoniakem (amoniakový pufr): H + + NH3 ↔ NH4 +
Hemoglobinový systém se tedy účastní dvou procesů:
Vazba protonů nahromaděných v důsledku metabolických procesů;
Protonace hydrogenuhličitanových iontů s následným uvolněním CO2
Systém hemoglobinového pufru lze považovat za jeden z nejdůležitějších článků transportu CO2 z tkání do plic.
Je třeba poznamenat, že udržování konstantního pH různých kapalných systémů těla není ovlivněno ani tak pufračními systémy, ale fungováním řady orgánů a systémů: plic, ledvin, střev, kůže atd.
Jedná se o schopnost pufrovacího systému působit proti změnám pH prostředí.
Nazývá se rozsah hodnot pH, nad a pod kterým přestává pufrační účinek nárazníková zóna.
Je rovnocenná pH = pK ± 1
Pufrovací kapacita (B) je vyjádřena jako počet molárních ekvivalentů silné kyseliny nebo zásady, které je nutné přidat do jednoho litru pufru, aby se pH posunulo o jednu.

B – vyrovnávací kapacita,
nЭ – množství molárního ekvivalentu silné kyseliny nebo zásady,
pHН – počáteční hodnota pH (před přidáním kyseliny nebo zásady)
pHК – konečná hodnota pH (po přidání kyseliny nebo zásady)
V praxi se vyrovnávací kapacita vypočítá pomocí vzorce:

V – objem kyseliny nebo zásady,
N – ekvivalentní koncentrace kyseliny nebo zásady,
Vfanoušek.– objem tlumivého roztoku,
Následující údaje naznačují, že působí proti změnám pH krve. Chcete-li posunout pH krve o jednu jednotku do alkalické oblasti, musíte do krve přidat 70krát více NaOH než do stejného objemu čisté vody. Pro změnu pH o jednu na kyselou oblast by mělo být do krve přidáno 320krát více kyseliny chlorovodíkové než do stejného objemu čisté vody.
Kapacita pufru závisí na koncentraci elektrolytů a poměru složek pufru. Roztoky s vyšší koncentrací složek a poměrem složek rovným jedné mají největší pufrační kapacitu.
Pufrovací kapacita arteriální krve je 25,3 mmol/l, venózní krve 24,3 mmol/l, sliny mají pufrační kapacitu a jsou určeny bikarbonátovým, fosfátovým a proteinovým systémem. Tlumicí kapacita slin se mění pod vlivem řady faktorů: sacharidová dieta snižuje tlumivou kapacitu slin, vysokoproteinová dieta ji zvyšuje. Výskyt zubního kazu je menší u jedinců s vysokou pufrovací kapacitou.
V lidském těle působí proteinové, hemoglobinové, fosfátové a bikarbonátové pufry.
Tlumivé systémy těla.
Tvoří 53 % kapacity vyrovnávací paměti a je reprezentován:
Bikarbonátový pufr je hlavním pufrovacím systémem krevní plazmy; je to systém rychlé reakce, protože je produktem jeho interakce s kyselinami CO2 – rychle se vylučuje plícemi. Kromě plazmy se tento pufrový systém nachází v červených krvinkách, intersticiální tekutině a ledvinové tkáni.
Mechanismus působení.
Pokud se kyseliny hromadí v krvi, množství HCO klesá3 – a dojde k reakci: NSO3 – + N + ↔ N2S3 ↔ N2O + CO2↑. Přebytek je odstraněn plícemi. Hodnota pH krve však zůstává konstantní, protože objem plicní ventilace se zvyšuje, což vede ke snížení objemu CO2
Se zvýšením alkality krve, koncentrace HCO3 – zvyšuje: N2S3 + OH – ↔ NSO3 – + N2O.
To vede k pomalejší ventilaci plic, takže CO2 se hromadí v těle a poměr pufrů zůstává nezměněn.
Představuje 35 % kapacity vyrovnávací paměti.
Hlavní pufrovací systém červených krvinek, který tvoří asi 75 % celkové pufrační kapacity krve. Účast hemoglobinu na regulaci pH krve je spojena s jeho úlohou v transportu kyslíku a CO2. Systém hemoglobinového pufru v krvi hraje významnou roli v několika fyziologických procesech najednou: dýchání, transport kyslíku v tkáních a udržování konstantního pH uvnitř červených krvinek a nakonec v krvi.
Je reprezentován dvěma slabými kyselinami – hemoglobinem a oxyhemoglobinem a jejich konjugovanými bázemi – hemoglobinátovými a oxyhemoglobinátovými ionty:
Oxyhemoglobin je silnější kyselina (pKа = 6,95) než hemoglobin (pKа = 8,2). Při pH = 7,25 (uvnitř červených krvinek) je oxyhemoglobin ionizován o 65 % a hemoglobin o 10 %, takže přidání kyslíku k hemoglobinu snižuje hodnotu pH krve, protože se tvoří silnější kyselina. Na druhou stranu, jak se z oxyhemoglobinu ve tkáních uvolňuje kyslík, hodnota pH krve se opět zvyšuje.
Tlumivé vlastnosti HHb jsou primárně dány možností interakce kyselino-reaktivních sloučenin s draselnou solí hemoglobinu za vzniku ekvivalentního množství odpovídající draselné soli kyseliny a volného hemoglobinu:
Výsledný hydrogenuhličitan (HCO3) vyrovnává množství příchozích H2S3pH se udržuje, když se potenciální molekuly H disociují2S3 a výsledné hemoglobinové kyseliny.
Takto se pH krve udržuje v normálních mezích i přes vstup obrovského množství CO do žilní krve2 a další kysele reaktivní metabolické produkty.
V kapilárách plic hemoglobin (HHb) absorbuje kyslík a mění se na HHbO2, což vede k určitému okyselení krve, vytěsnění určitého množství H2S3 z bikarbonátů a snížení alkalické rezervy krve a v tkáních ji odevzdává a absorbuje CO2.
Kromě toho je hemoglobinový pufr komplexní protein a působí jako proteinový pufr.
Tvoří 5 % kapacity vyrovnávací paměti. Obsaženo jak v krvi, tak v buněčné tekutině jiných tkání, zejména ledvin. V buňkách je reprezentován K solemi2NRA4 a KN2RO4a v krevní plazmě a mezibuněčné tekutině Na2HPO4 a NaH2PO4. Funguje hlavně v plazmě a zahrnuje: dihydrogenfosfátový iont H2RO4 – a hydrogenfosforečnanový ion HPO4 2-.
Poměr [HPO4 2-]/[H2PO4 – ] v krevní plazmě (při pH = 7,4) se rovná 4:1. V důsledku toho má tento systém pufrační kapacitu pro kyselinu větší než pro zásadu.
Například při zvýšení koncentrace kationtů H + v intracelulární tekutině, například v důsledku zpracování masité potravy, jsou neutralizovány ionty HPO4 2-:
Vzniklý nadbytek dihydrogenfosforečnanu je vylučován ledvinami, což vede ke snížení pH moči.
Když se koncentrace zásad v těle zvýší, například při konzumaci rostlinné stravy, jsou neutralizovány H ionty2RO4 1-:
Vzniklý nadbytek hydrogenfosforečnanu je vylučován ledvinami a zvyšuje se pH moči.
Vylučování určitých složek fosfátového pufrového systému močí v závislosti na zpracovávané potravině vysvětluje široký rozsah hodnot pH moči – od 4,8 do 7,5. Fosfátový pufrovací systém krve se vyznačuje nižší pufrovací kapacitou než bikarbonátový systém v důsledku nízké koncentrace krevních složek. Tento systém však hraje rozhodující roli nejen v moči, ale i v dalších biologických prostředích – v buňce, ve šťávách trávicích žláz, v moči.
Tvoří 5 % kapacity vyrovnávací paměti. Skládá se z bílkovinné kyseliny a její soli tvořené silnou zásadou.
Pt – COOH – protein-kyselina
Pt – COONa – protein-sůl
Když se v těle tvoří silné kyseliny, interagují s proteinovou solí. Tím vzniká ekvivalentní množství proteinové kyseliny: HC1 + Pt-COONa ↔ Pt-COOH + NaCl. Podle zřeďovacího zákona W. Ostwalda zvýšení koncentrace slabého elektrolytu snižuje jeho disociaci, pH zůstává prakticky nezměněno.
Jak alkalické potraviny přibývají, interagují s
Pt-COOH: NaOH + Pt-COOH ↔ Pt-COONa + H2O
Množství kyseliny klesá. Koncentrace H+ iontů se však zvyšuje v důsledku potenciální kyselosti proteinové kyseliny. pH se tedy prakticky nemění.
Protein je amfoterní elektrolyt, a proto vykazuje svůj vlastní pufrovací účinek.
Podívejme se na interakci nárazníkových systémů v těle ve fázích:
Během výměny plynů v plicích vstupuje kyslík do červených krvinek, kde probíhá reakce:
Při pohybu krve do periferních částí oběhového systému se z ionizované formy HbO uvolňuje kyslík2 –
V tomto případě se krev mění z arteriální na venózní. Kyslík uvolněný ve tkáních se spotřebuje na oxidaci různých substrátů, což má za následek tvorbu CO2, z nichž většina vstupuje do erytrocytů.
V erytrocytech v přítomnosti karboanhydrázy dochází významnou rychlostí k následující reakci:
Výsledné přebytečné protony se vážou na hemoglobinátové ionty:
Vazbou protonů se posouvá rovnováha reakce stupně (3) doprava, v důsledku toho se zvyšuje koncentrace hydrogenuhličitanových iontů a ty difundují membránou do plazmatu. V důsledku protidifúze iontů lišících se acidobazickými vlastnostmi (chloridový iont je protolyticky neaktivní, hydrogenuhličitanový iont je v tělesných podmínkách zásadou) hydrokarbonát-chloridový posun. To vysvětluje kyselejší reakci prostředí v erytrocytech (pH = 7,25) ve srovnání s plazmou (pH = 7,4).
Bikarbonátové ionty vstupující do plazmy neutralizují přebytečné protony, které se tam hromadí v důsledku metabolických procesů:
Výsledný CO2 interaguje se složkami proteinového pufrovacího systému:
S2 + Рt-NH2 ↔ Pt-NHCOOH ↔ H + + Pt-NHCOO –
Přebytečné protony jsou neutralizovány fosfátovým pufrem:
Po návratu krve do plic se v ní zvyšuje koncentrace oxyhemoglobinu (1. stupeň), který reaguje s hydrogenuhličitanovými ionty, které nedifundovaly do plazmy:
Výsledný CO2 se vylučuje plícemi. V důsledku poklesu koncentrace HCO3 – jsou pozorovány ionty v této části krevního řečiště, jejich difúze do erytrocytů a difúze chloridových iontů v opačném směru.
Ledviny také hromadí přebytečné protony v důsledku reakce:
který je neutralizován hydrofosfátovými ionty a amoniakem (amoniakový pufr): H + + NH3 ↔ NH4 +
Hemoglobinový systém se tedy účastní dvou procesů:
Vazba protonů nahromaděných v důsledku metabolických procesů;
Protonace hydrogenuhličitanových iontů s následným uvolněním CO2
Systém hemoglobinového pufru lze považovat za jeden z nejdůležitějších článků transportu CO2 z tkání do plic.
Je třeba poznamenat, že udržování konstantního pH různých kapalných systémů těla není ovlivněno ani tak pufračními systémy, ale fungováním řady orgánů a systémů: plic, ledvin, střev, kůže atd.
















